Микроэволюция инвазионных видов
Успешному прогнозированию результатов инвазионного процесса препятствуют две проблемы:
- Невозможность предугадать вероятных «захватчиков»;
- Степень нарушенности естественной растительности не играет такой определяющей роли, которая отводилась ей ранее.
Если бы можно было точно предсказать, что конкретный вид или вид, обладающий определённым набором биологических признаков, будет самостоятельно завоёвывать новые территории, задача состояла бы в усилении карантинной службы. Однако проблема осложняется тем, что заносные виды могут долгое время произрастать на ограниченной территории, и лишь затем происходит взрывное расширение их ареала. К сожалению, моделей, прогнозирующих вторжение заносных видов, не так много, и затрагивают они, в лучшем случае, только узкий диапазон организмов. Между тем ботанические сады и садоводческие фирмы постоянно вводят в культуру как новые дикорастущие виды, так и виды, успешно культивируемые в других регионах, тем самым приводя в действие пусковой механизм микроэволюции. Начальная популяция интродуцированного вида неустанно расширяется (благодаря усилиям по его размножению), но имеется возможность постоянного возникновения и новых центров (например, за счёт введения в культуру других сортов того же вида на смежной территории). Интродукция растений — процесс, как правило, многократно повторяющийся; чаще всего он дублируется в разных точках континентов или стран.
Что же происходит после введения вида в культуру? Большинство интродуцированных растений произрастают в ботанических садах лишь благодаря действиям человека. Однако некоторая часть видов доходит до стадии колонизации, то есть способности размножаться самосевом, а половина из этой части достигает стадии натурализации, то есть «сбегает из культуры» в естественные ценозы. Таким образом, ботанические сады являются как бы резервуарами, подпитывающими естественные ценозы новыми видами, а конкретные флоры постоянно обогащаются «сбежавшими экспонатами».
При изучении истории интродукции широко натурализовавшихся в России в настоящее время растений установлено, что все из исследованных однолетних видов являются беженцами из ботанических садов [Виноградова, 1992]. Так, мелколепестник канадский Conyza canadensis в 1646 году был завезён в ботанический сад Нюрнберга как редкостное заморское растение. В XVIII веке он был обнаружен в Южной Германии уже в качестве адвентивного вида. Эхиноцистис шиповатый Echinocystis lobata сбежал из культуры в ботаническом саду в Румынии в 1904 году. Из ботанического сада Потсдама в 1896 году сбежала североамериканская Bidens frondosa. Недотрогу мелкоцветковую Impatiens paroiflora в конце XIX века выращивали в ботанических садах Львова, Санкт-Петербурга, Тарту, Риги и Пярну, откуда она и сбежала через 50 лет. Chamomilla suaoeolens сбегала дважды: из ботанического сада Гельсингфорса (Финляндия) в 1849 году и из Краковского ботанического сада в 1886 году. Южноамериканскую галинзогу мелкоцветковую Galinsoga paroiflora культивировали в 1785 году в Парижском ботаническом саду, откуда и ведут своё происхождение большинство европейских популяций этого вида. Перечень беглецов можно пополнить и такими широко распространёнными видами, как Amaranthus retroflexus, Veronica persica, Cyclachaena xanthiifolia, которые через ботанические сады разбрелись по всей Европе и стали злостными сорняками.
По счастью, не все заносные виды способны к натурализации. Подсчитано, что только 10% заносных видов успешно доходят до стадии колонизации и лишь 5% — натурализации [Considine, 1986]. К тому же иммиграционный период (лаг-фаза) длится обычно довольно долго. На примере ряда однолетних растений нами установлено, что между первым появлением вида на новой территории и его внедрением в естественные ценозы может пройти до 100 смен поколений. Epilobinm adenocaidon, имеющий естественный ареал в Северной Америке, собран впервые в Британии в 1891 году и произрастал там в течение полувека, не расширяя ареала. В Финляндии этот вид стал бурно расширять ареал через 20 лет после первого появления. В Европейской России появился не позднее 1909 года, но стал обычным растением только через 50 лет. Бурное расселение Bidens frondosa началось лишь после Второй мировой войны — через 50 лет. Гималайский вид Impatiens glandidifera культивировался в России с конца XIX в., а бурная его натурализация началась лишь через 60-70 лет. Среднеазиатская I. paroiflora появилась в Западной Европе в 1830-х годах, дичать стала через 10-15 лет, а стремительное расширение её ареала и натурализация начались лишь через 120 лет. Galinsoga parvilflora интродуцирована в Англию в 1796 году, найдена там вне ботанического сада через 13 лет, а массовое ее распространение началось только через 125 лет! Североамериканский Amaranthus albus впервые был отмечен в Италии в 1723 году, в Средней Европе появился в конце XIX века, натурализовался только к 1960-м годам, то есть через 250 лет! За этот период в первоначальной генетически обеднённой инвазионной популяции происходит накопление генетической изменчивости.
Какими же путями происходит изменение генофонда интродуцированных растений в ботанических садах? Здесь имеют место все четыре основных фактора эволюции, два из которых (мутационный процесс и поток генов) создают изменчивость, а два (отбор и дрейф генов) регулируют эту изменчивость.
Cравнительная оценка факторов микроэволюции в естественных фитоценозах и в ботанических садах
| Естественные фитоценозы | Ботанические сады |
|---|---|
| Мутационный процесс | |
| Преимущество выживания доминантной аллели А1 | В новых условиях может получить преимущество рецессивная аллель а2 |
| Рецессивная аллель а2 встречается редко, в основном в гетерозиготном состоянии | Искусственное сохранение и размножение рецессивной аллели а2 |
| Поток генов | |
| Скорость потока генов невелика | Скорость потока генов возрастает. Увеличивается вероятность появления рецессивной аллели а2 за счёт многократной интродукции |
| Отбор | |
| Только естественный отбор | Естественный и искусственный отбор |
| Идёт стабилизирующий отбор, дающий преимущество генотипам, не отклоняющимся от нормы | В новых условиях среды возрастает роль ведущего отбора |
| Возникшие макромутанты обычно нежизнеспособны | Макромутанты размножаются искусственно |
| Дрейф генов | |
| Минимален, так как в многочисленной популяции есть возможность восполнить потерю утраченного генотипа | Низкая численность особей в интродукционной популяции. Кража или повреждение ведут к утрате генотипа |
Мутационный процесс в ботанических садах поддерживается благодаря искусственному сохранению растений с рецессивной аллелью a2, нередко представляющих интерес с декоративной точки зрения. В новых условиях среды эти растения могут даже получить преимущество выживания. Напротив, в естественных устойчивых популяциях преимущество выживания имеет доминантная аллель А1, тогда как рецессивная а2 встречается крайне редко, в основном в гетерозиготном состоянии.
Скорость потока генов в ботанических садах намного выше, чем в естественных популяциях, поскольку многократная интродукция декоративных видов из разных источников (из разных природных популяций или разных ботанических садов) увеличивает вероятность многократного поступления рецессивной аллели а2 в интродукционные популяции. Процесс этот усугубляется ещё и тем, что в последнее время в России в связи со значительным сокращением экспедиционных поездок ботанические сады стали друг для друга основными поставщиками исходного интродукционного материала. В природных условиях возникшие макромутанты (в основном рецессивные) обычно нежизнеспособны и, как правило, погибают. В ботанических же садах, применяя искусственное вегетативное размножение растений, макромутанты можно сохранять сколь угодно долго. Этим способом в конечном счёте созданы практически все сорта широко распространённых вегетативно размножающихся декоративных растений: розы, тюльпаны, сирень и так далее.
Значительные сдвиги частот аллелей A1 и A2 могут происходить в интродукционных популяциях случайным образом. Действие этого чисто случайного компонента в изменении генных частот, известного под названием дрейфа генов, в естественных популяциях практически сведено к нулю, поскольку в многочисленной популяции всегда есть возможность восполнить потерю утраченного генотипа. Однако в ботанических садах из-за низкой численности культивируемых особей любое случайное повреждение или кража десятка растений может привести к полной утрате какого-либо гена в интродукционной популяции. Этот фактор сильно влияет на генофонд декоративных растений, поскольку именно они чаще всего становятся объектами подобного варварского отношения со стороны посетителей ботанических садов.
Таким образом, в интродукционных популяциях, по сравнению с природными, все факторы эволюционного процесса (кроме естественного отбора) действуют интенсивнее и, помимо того, прибавляются ещё два мощнейших фактора — искусственный отбор и искусственная гибридизация. Факторы, создающие изменчивость, дают начало процессу микроэволюции, а факторы, регулирующие изменчивость, продолжают этот процесс, что приводит к установлению новых частот вариантов. Генофонд интродукционных популяций столь сильно преобразуется, что можно говорить о наличии процесса видообразования в ботанических садах. К сожалению, работ по сравнению генофонда естественных и интродукционных (переживших несколько смен поколений) популяций одного и того же вида слишком мало. Однако многочисленные примеры значительного расширения ареала «сбежавших из культуры» видов являются неоспоримым свидетельством микроэволюции растений во вторичном ареале.
Из всего вышеизложенного вытекают три проблемы. Во-первых, встаёт вопрос: можно ли считать, что интродукционные (особенно натурализовавшиеся) и естественные популяции относятся к одному и тому же виду? Эта проблема особенно остро касается агамоспермных видов. Если последовательно применять критерии выделения видов, можно столкнуться со множеством трудностей. Для современной систематики вид есть понятие морфолого-эколого-географическое, а по всем этим признакам дикорастущие и культивируемые особи сильно различаются. Так, широко распространённая в России черноплодная арония, отличающаяся от родоначального вида Aronia melanocarpa по числу хромосом, способу опыления, количеству и размерам цветков и плодов, форме листовой пластинки, зимостойкости, была описана в качестве нового вида Aronia mitschurinii [Скворцов, Майтулина, 1982]. Во-вторых, установление новых частот аллелей в интродукционных популяциях является важнейшим препятствием для успешности процесса репатриации видов в естественные условия произрастания. Требуется такое же (если не большее) число смен поколений для возврата генофонда в исходное состояние, независимо от того, изменились или нет условия среды в «родоначальном» местообитании репатриируемого вида. И в-третьих, «растением-захватчиком» потенциально может стать любой интродуцированный вид. Следовательно, анализ генетических ресурсов ботанических садов на предмет выявления видов, которые размножаются самосевом и в ближайшем будущем могут превратиться в сорняки, должен стать важной задачей на ближайшее десятилетие.
Внутривидовая изменчивость заносных видов выявлена у многих растений, имеющих обширный вторичный ареал: у Zizania aquatica отмечена изменчивость количественных признаков в северном Саскачеване, куда она занесена была в 1935 году. [Peden, 1982]; Echium vulgare, занесённый из Старого Света в Северную Америку в начале XIX века, обладает настолько высокой амплитудой изменчивости, что некоторые авторы описывают его разновидности [Gleason, 1963]; мультивариантный анализ по 64 морфологическим признакам, проведённый в популяциях Bromus japonicus в штате Вашингтон, показал, что экземпляры из мезофитных местообитаний имеют более значительную межпопуляционную изменчивость, чем из ксерофитных [Heidel et al., 1982].
Широкую вариабельность проявляют виды, интродуцированные в Австралию в XIX веке — Trifolium glomeratum [Woodward, Morley, 1974], три вида рода Avena [Whalley, Burfitt, 1972] и другие.
Изучение внутривидовой изменчивости интродуцированного в Северной Америке Avena barbata на уровне генома [Allard, 1975] позволило выделить у этого вида 35 вариаций генома, три из которых распространены довольно широко.
Изучение внутривидовой изменчивости Trifolium hirtum показало, что разновидность Wilton, завезённая в Калифорнию, претерпела значительные эволюционные преобразования, причём уже в самых ранних по времени пастбищных посевах. Межпопуляционные различия отмечены и по морфологическим, и по количественным, и по ферментативным признакам, однако большинство генетических изменений проявлялось в виде изменения частот аллелей, а не в виде появления новых аллелей. У калифорнийских популяций отсутствуют две аминокислоты, имеющиеся в родоначальных турецких растениях. Разновидности были более гомозиготными, чем естественные популяции, но они также проявили большое генетическое разнообразие. Внутрипопуляционная изменчивость отмечена для количественных признаков и времени цветения. Авторы сделали вывод, что расширение ареала вида на новой территории обеспечивают быстрые микроэволюционные изменения, но не путём изоляции и дрейфа, а, более вероятно, силами отбора [Martins, Jain, 1980].
При изучении генетической структуры 19 дикорастущих и культивируемых популяций Pennisetum typhoides было показано [Pilate-Andre et al., 1987], что отдельные локусы различаются по уровню полиморфизма аллелей. Уровень изменчивости дикорастущих популяций везде выше, чем культивируемых, причём наблюдается уменьшение градиента изменчивости популяций с востока на запад африканского континента.
При изучении изменчивости аллозимов у трёх видов Aeschynomene показано [Carilli, Fairbrothers, 1988], что заносный сорняк A. indica имеет большую, по сравнению с аборигенным видом A. virginica, вариабельность.
Повышение генетической изменчивости в ходе колонизации растениями новых местообитаний отмечено и у других интродуцированных видов: Ricinus communis [Harland, 1947] и Avena fatua [Jain, Marshall, 1967].
Эволюция генетической пластичности наглядно изучена на примере Sarracenia purpurea. Первая популяция вида возникла от посаженного на сфагновом островке в штате Огайо одного экземпляра. С помощью метода электрофореза аллозимов была изучена изменчивость популяции — «основателя» и 10 других популяций в США. В популяции на сфагновом островке изменчивость была самой низкой — найден лишь один полиморфный локус (в некоторых популяциях половина локусов была полиморфной), а гетерозиготность на особь на 50% ниже, чем средняя по виду [Schwaegerle, Schaal, 1979].
По данным электрофореза ферментов, уровни генетической изменчивости у Helianthns bolanderi ssp. exilis и сорных ssp. bolanderi и H. annuus существенно не различаются [Rieseberg et al., 1988]. Различия наблюдаются в распределении изменчивости: у Н. bolanderi ssp. exilis — в пределах популяции, а у сорных таксонов — между популяциями.
При изучении изменчивости 10 ферментных систем методом электрофореза у двух интродуцированных в Америку видов клевера установлено, что у обоих видов изменчивы 8 локусов из 15 (у Trifolium hybridum в среднем 1,73 аллели на локус, а у T. pratense — 1,93 аллели на локус). Межпопуляционная изменчивость у обоих видов составляет в среднем 10%, а внутрипопуляционная изменчивость довольно высокая [Hickey, Vincent, Guttman, 1991].
Определение среднего значения признаков, меры их пластичности, характера фенотипической пластичности для видов в пяти родах и подсчёт расхождений между видами по каждому из этих измерений показало, что фенотипическая пластичность признаков может эволюировать независимо от самих признаков и что дивергенция видов по пластичности признаков вполне реальна. Растения в одной популяции могут различаться по фенотипической пластичности; возможен отбор по этому признаку [Schlichting, Levin, 1986].
Интереснейшая работа проведена с Lupinus succidentus — американским однолетником, который в Калифорнии стал поселяться на рудеральных местах. Сравнивали генные частоты в естественных и рудеральных популяциях этого вида и изучали полиморфизм в четырёх локусах. Три локуса (ВВ, РР и DD) отвечают за окраску цветков, а локус SS определяет рисунок семенной кожуры. Большинство популяций L. succidentus оказались мономорфными по локусам ВВ, РР и DD. Лишь в одном из каньонов найдены популяции, полиморфные по локусу D/d, здесь наблюдалась клинальная изменчивость — от основания к вершине каньона уменьшалось количество голубых цветов. Локус S/ s оказался полиморфным в подавляющем большинстве популяций, но чаще это наблюдалось в популяциях, растущих в нарушенных местообитаниях. Полиморфизм в локусах D/d, B/b и Р/р найден только в нарушенных местообитаниях. Таким образом, небольшие популяции, начинающие колонизировать новые местообитания, не только не являются генетически однородными, но, наоборот, имеют большую генетическую изменчивость, чем естественные популяции.
Данные о лабильности генетического аппарата видов-колонизаторов имеются в ряде других работ [Kornas, 1982; Oka, 1983]. Однако различия между таксонами по пластичности отдельных признаков изучены недостаточно.
Экспериментальное изучение внутривидовой изменчивости инвазионных растений впервые начал Дохальд, который отметил у адвентивного в Австралии Trifolium subterraneum существование нескольких рас, различающихся по срокам созревания при выращивании их в однородных условиях. Позднее изучение внутривидовой изменчивости этого же вида позволило выделить в Австралии более 130 его рас, различающихся не только по морфологическим признакам, но и по устойчивости к болезням [Morley, Katznelson, 1965; Kellock et al., 1978].
Наиболее детально изучена вариабельность Chondrilla juncea — облигатного апомикта, появившегося в Австралии в 1917 году Исследованиями McVean [1966] установлено, что в Австралийском регионе этот вид морфологически единообразен. Но уже через 6 лет обнаружены три формы Ch. juncea, различающиеся по морфологии соцветия, характеру плодов и форме розеточных листьев [Hull, Groves, 1973]. По меньшей мере, в двух генерациях растений, выращенных в однородных условиях, эти морфологические различия сохранялись. Формы различались также по продолжительности префлорального периода и способности к вегетативному размножению. Среди растений формы А отмечена клинальная изменчивость префлорального периода, который увеличивается с запада на восток.
Все три формы (А, В и С) произрастают вместе на площади ~3,3 миллиона га. Вокруг этого участка имеется концентрический ареал, в котором встречаются только формы А и С. На внешней стороне этого второго ареала найдены растения только формы А, которые широко распространены и встречаются в Юго-Восточной Австралии в районах с разным климатом и типом почв. Ареалы всех трёх форм продолжают увеличиваться.
Применение электрофоретического метода позволило установить, что 8 энзимов из исследованных 14 дают для каждой из трёх форм различные изозимные пятна [Burdon et al.,1980]. Дальнейшие исследования касались влияния трёх популяций клеща Aceria chondrillae на три формы Ch. jnncea [Cullen, Moore, 1983]. Выяснилось, что первая популяция клеща поражает только форму А, а две другие — только форму В.
У двух близких видов Hordenm leporinnm и H. glaucum, естественно произрастающих в Европе, Западной Азии и Северной Америке, во вторичном ареале в Австралии (где комплекс этих видов появился в 1840-1850-х годах), отмечена межпопуляционная изменчивость времени вступления в фазу цветения [Cocks et al., 1976]. Признак этот находится в коррелятивной зависимости от длины вегетационного периода в пункте сбора семян, причём у H. leporinum коррелятивная связь была более тесной, чем у H. glaucum.
Выявлен диапазон внутривидовой изменчивости Bromus mollis — адвентивного вида североамериканской флоры [Allard, 1965]: количество побегов и высота растений изменяются незначительно, а период развития от всходов до колошения тем меньше, чем южнее произрастает популяция.
Имеются также экспериментальные данные об отличиях между популяциями различного географического происхождения адвентивного в Америке Rumex crispus по требованиям к яровизации и фотопериодизму [Hume, Cavers, 1983].
Таким образом, в более или менее заметных масштабах изучение внутривидовой изменчивости инвазионных видов ведётся лишь последние 30 лет. За это время были получены данные об изменчивости не более чем сотни видов-колонизаторов, причём экспериментальными методами изучено B 50 из них.
Наши исследования, проведённые в Главном ботаническом саду им. Н. В. Цицина РАН [Виноградова, 1992], охватили 16 заносных видов, принадлежащих к различным жизненным формам, с различной биологией размножения, в разное время и разным путём занесённые в нашу страну: Anisantha tectorum, Acorns calamus, Amaranthus albus, Ribes aureum, Aronia mitschurinii, Acer negundo, Impatiens glandidifera, I. parviflora, Epilobium adenocaidon, E. pseudorubescens, Echinocystis lobata, Bidens frondosa, Conyza canadensis, Chamomilla suaveolens, Galinsoga paroiflora, G. quadriradiata. Общим для них было только то, что они претерпевают процесс активной натурализации и расширения ареала.
Тем не менее следует понимать, по сравнению с громадным числом заносных видов, встречающихся в России, это капля в море. Требуются вовлечение в исследования гораздо более широкого набора объектов и изучение их по сходной методике. В ходе исследования должны быть зафиксированы достоверные микроэволюционные сдвиги, произошедшие в натурализующихся видах за более или менее точно определимый срок. Под микроэволюционным сдвигом мы понимаем такое изменение генофонда популяции, которое приводит к достаточно ясному изменению адаптивных свойств этой популяции. Появление отдельных мутаций или каких-либо индивидуальных отклонений, адаптивный смысл которых неясен, в счёт не идёт. В связи с этим считаем целесообразным привести, хотя бы схематично, методологию исследования внутривидовой изменчивости и микроэволюции заносных видов:
- Изучение истории интродукции и составление карты естественного и вторичного ареалов вида по литературным источникам и гербарным образцам, что позволит определить продолжительность периода эволюционной дифференциации;
- Выявление диапазона модификационной изменчивости морфологических признаков в природных условиях путём обследования естественных и инвазионных популяций изучаемых видов, причём особое внимание уделяя признакам, имеющим диагностическое значение;
- Изучение биологических признаков семян различного географического происхождения посредством проращивания семян в лабораторных при различных температурных режимах и сроках стратификации, а в условиях питомника — при различных сроках посева: подзимнем, весеннем и летнем;
- Выращивание растений в однородных почвенно-климатических условиях из семян различного географического происхождения (создание интродукционных популяций). Сбор семенного материала проводится вдоль широтного профиля вторичного ареала изучаемых видов — от крайних северных до крайних южных точек произрастания вида на территории России. Для более полного изучения полиморфизма растений можно использовать также семена, присланные по делектусам. Для определения эндогенной изменчивости семена собирают с каждого экземпляра отдельно. Внутрипопуляционную изменчивость оценивают в интродукционных популяциях, сформированных из семян, собранных в 4-6 микропопуляциях с 10-15 растений в каждой, в двух повторностях. Выращивание образцов всех сравниваемых популяций в однородных условиях питомника позволяет оценить действительно генотипы, а не вариации фенотипов;
- Изучение в интродукционных популяциях темпов и продолжительности роста, фенологии и соответствующих фенофазам морфологических изменений, способа опыления; для некоторых видов необходимо изучение вопроса об озимости/яровости, а также фотопериодической реакции сравниваемых образцов;
- Сравнение диапазона и характера изменчивости изучаемых видов в первичном и вторичном ареалах;
- Выявление различий изучаемых видов в естественном и вторичном ареалах молекулярно-генетическими методами;
- В очень редких случаях сравнение конкурентоспособности близкородственных аборигенного и заносного видов.
Целью подобных исследований должны стать выявление основных закономерностей микроэволюции, происходящих при инвазии растений, и определение факторов, влияющих на процессы микроэволюции, а также диагностика признаков, способствующих широкой натурализации видов-колонизаторов, и установление зависимости процессов микроэволюции от способа размножения, жизненной формы, отношения к длине дня и других биологических признаков.
Результаты изучения амфимиктных видов
Acer negundo появился в России почти двести лет назад, но на большей части ареала он произрастает лишь с конца XIX — начала XX века, а первые упоминания о внедрении клёна в аборигенную флору и размножения его самосевом относятся к 1920-м годам. Учитывая, что клён вступает в стадию плодоношения на 6-7-м году жизни, можно утверждать, что эволюция приспособительных признаков затронула лишь 9-11 поколений вида.
В естественном ареале у A. negundo наблюдается клинальная изменчивость количественных признаков плодов: от севера к югу происходит постепенное уменьшение длины, ширины и массы крылатки. Длина крылаток северного происхождения превышает длину крылаток южного происхождения в 1,5 раза, а масса крылаток — в 2 раза [Williams, Winstead, 1972; Виноградова, 2006]. Во вторичном ареале изменчивость плодов изучена в инвазионных популяциях в Ашхабаде, Астрахани, Москве и Вологде. Внутрипопуляционная изменчивость оказалась довольно высокой: длина крылатки варьировала у ашхабадских образцов от 3,7 до 4,6 см, у московских — от 3,3 до 4,5 см, а у астраханских и вологодских — от 2,9 до 4,6 см (коэффициент вариации 20-25%). Наибольшая амплитуда изменчивости (коэффициент вариации 30-36%) отмечена для средней массы крылаток, которая варьировала от 20 до 88 мг. Однако по среднему значению длины, ширины и массы крылаток исследованные инвазионные популяции практически не различались. Таким образом, во вторичном ареале внутривидовая изменчивость количественных признаков плодов A. negundo не носит чётко выраженного географического характера и является в основном внутрипопуляционной.

Внутривидовая изменчивость крылаток A. negundo во вторичном ареале
В лабораторных условиях семена различного географического происхождения проявили сходство по требованиям к определённой температуре прорастания и сроку стратификации, однако семена A. negundo из южных частей вторичного ареала обладали меньшим периодом покоя, чем семена из более северных районов.
Интродукционная популяция была заложена из семян, собранных в Ашхабаде, Астрахани, Москве и Вологде. Семена московского происхождения имели наиболее высокую всхожесть — 69%; всхожесть астраханских семян — 40%. Семена, собранные в крайних точках ареала — Вологде и Ашхабаде, имели очень низкую всхожесть (вологодские — 23%, ашхабадские — 26%), а также низкую энергию прорастания, которая вычислялась за первые 15 дней.
Установлено наличие клинальной изменчивости ритма развития однолетних сеянцев. Чем севернее находится пункт сбора семян, тем раньше распускается соответствующая пара листьев у выросших из них растений; различие между образцами вологодского и ашхабадского происхождения составляет 2 недели. Сеянцы московской репродукции в основном закончили свой рост в первой декаде сентября, вологодской и астраханской репродукций — на неделю позже, а ашхабадской репродукции — на месяц позднее. В связи с продолжительным периодом роста даже к концу ноября хорошо сформированной верхушечной почки не было у 44% сеянцев московской репродукции, у 71% сеянцев вологодской репродукции, у 77% сеянцев астраханской и у 92% сеянцев ашхабадской семенных репродукций.
Однолетние сеянцы в большой степени пострадали от мороза в суровую зиму 1978/79 годов. Полностью вымерзли 6% сеянцев ашхабадской репродукции, остальные 94% сильно обмёрзли (в среднем на 5-6 междоузлий, то есть на 24 см). Вымерзли также 2% сеянцев вологодской и 1% сеянцев астраханской репродукций, соответственно 12 и 3% из них существенно обмёрзли. Среди сеянцев московского происхождения полностью вымерзших экземпляров не было, 23% сеянцев совершенно не пострадали от морозов, а у остальных подмёрзли 1-2 междоузлия (6-7 см). К началу второго вегетационного периода средняя высота растений московского происхождения в 3 раза превышала высоту растений ашхабадского происхождения ив 1,5 раза — высоту растений вологодского и астраханского происхождения.
За три периода вегетации наибольшим приростом по высоте, диаметру и биомассе отличались деревца местной (московской) семенной репродукции. У них отмечены также наименьшая продолжительность периода роста и наибольшая морозостойкость. К концу 3-го года жизни деревья московского происхождения были в 1,5 раза выше деревьев ашхабадского происхождения; последние к тому же из-за ежегодного обмерзания побегов имели кустообразную форму.
Таким образом, за 9-11 смен поколений у A. negundo уже выработалась генетически закреплённая клинальная географическая изменчивость некоторых биологических признаков: продолжительности периода покоя семян, морозостойкости деревьев, прироста по диаметру стволика и по биомассе растений, а также ритма их развития.
Impatiens glandulifera в качестве декоративного растения выращивали в садах Европы с середины XIX века, в Северной Америке — с начала XX столетия. Первые случаи дичания отмечены через 20 смен поколений, внедрение в естественные ценозы началось через 60, а бурная экспансия — через 100 смен поколений.
Интродукционная популяция была заложена при осеннем посеве семян, собранных в городах Санкт-Петербург, г. Крестцы Новгородской области, Вологда, Москва, Львов, Дубинки (Прикарпатье), Мукачево (Закарпатье).
Установлено наличие клинальной изменчивости ритма развития недотроги: чем севернее находится пункт сбора семян, тем быстрее развиваются растения. В интродукционной популяции растения из семян закарпатского происхождения вступают в генеративный период развития на 5 недель позднее, чем вологодские, причём 2/3 экземпляров не успевают даже вступить в стадию плодоношения, погибнув от первых заморозков ещё в цвету. Все растения северных образцов полностью проходят жизненный цикл и заканчивают его в третьей декаде сентября ещё до заморозков.
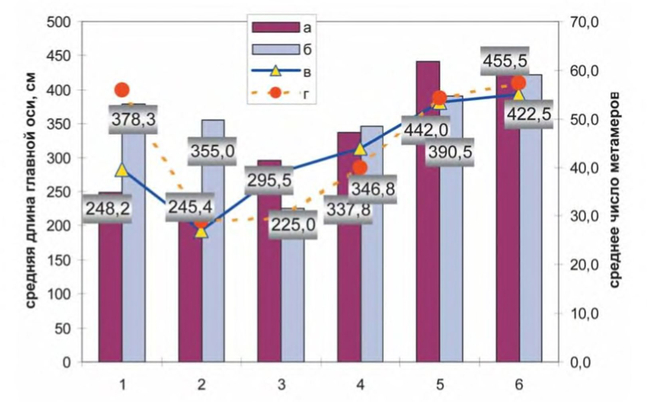
Отмечена клинальная изменчивость прироста по высоте и по биомассе растений: от севера к югу у сравниваемых образцов наблюдается достоверное увеличение средней высоты главного побега, среднего числа метамеров на главном побеге, биомассы надземной части, а также продолжительности периода роста растений. Северные образцы раньше закончили рост, и к концу вегетационного периода высота их была невелика: особи крестцовской репродукции в среднем достигли высоты 63 см и имели 10 метамеров, петербургской — 72 см (12 метамеров), московской — 90 см (10 метамеров) и вологодской — 107 см (12 метамеров). Южные образцы росли дольше, и к концу вегетации Львовские экземпляры имели среднюю высоту 122 см (13 метамеров), а мукачёвские — 111 см (13 метамеров).
Таким образом, за 150 смен поколений у I. glandidifera во вторичном ареале выработалась генетически закреплённая географическая изменчивость ритма роста и развития растений.
Экспериментально установлено, что в результате процесса адаптации и свободного переопыления растений в интродукционной популяции, в ряду последующих поколений микроэволюция адаптивных признаков идёт в сторону нивелирования различий между географическими образцами и сближением их с московскими экземплярами. Одновременное вступление в одну и ту же фенофазу и одинаковый темп роста наблюдались уже у растений 5-го поколения.

Феноритм I.glandulifera в ряду последовательных поколений (P, F1, F2, F3, F4)
1. Санкт-Петербург; 2. Крестцы; 3. Вологда; 4. Москва; 5. Львов; 6. Дубинки (Прикарпатье); 7. Мукачево (Закарпатье). а — вегетативный период; б — цветение; в — семеношение.
Изучено влияние длины дня на рост и фенологию образцов различного географического происхождения. Для этого сравнивали темпы роста и феноритмы растений, которые на стадии проростков в течение трёх недель росли при искусственном коротком дне (9 часов) с контрольными растениями, произраставшими в течение того же периода при естественной длине дня.
Особи вологодской, крестцовской и львовской репродукций, росшие при коротком дне, вступили в соответствующие фенофазы на 3 недели позднее, чем контрольные растения. У московского образца опытные растения вступили в генеративный период развития на неделю позднее контрольных. У дальневосточного образца опытные растения зацвели практически одновременно с контрольными.
У московского и владивостокского образцов влияние длины дня на высоту растений и на число метамеров достоверно не сказалось. У образцов крестцовского, вологодского и львовского происхождения, напротив, отмечался более медленный рост опытных растений.
Следовательно, в европейской части вторичного ареала недотрога железконосная является длиннодневным растением, а в дальневосточной части — растением, нейтральным по отношению к длине дня. В качестве гипотезы можно выдвинуть предположение, что длительное произрастание I. glandidifera в Европе до начала его стремительной натурализации связано с тем, что это короткодневное в естественном ареале растение не было способно эффективно обсеменяться в новых условиях. Однако, как только в результате рекомбинации генов появились экземпляры, способные полностью завершить жизненный цикл в условиях длинного дня, началось продвижение именно этой длиннодневной формы на север и стремительное расширение ареала недотроги. Наша гипотеза противоречит выводам английских учёных [Willis, Hulme, 2002] о ведущей роли температуры в комплексе факторов, лимитирующих расширение вторичного ареала недотроги. Нам она, однако, кажется предпочтительнее, поскольку в естественном ареале I. glandulifera достаточно холодостойка — ведь она растёт в горах до 3 500 м над уровнем моря!
Echinocystis lobata имеет в Евразии два изолированных очага расселения — Центральная Европа, где вид появился в начале XX века, и Дальний Восток, куда он был занесён непосредственно из Северной Америки в 1920-е годы. Бурное, взрывное расселение E. lobata по центральным районам Европейской России началось после 1960 года. В 1970-х годах он завоевал уже обширную территорию и вошёл в естественные ценозы Средней России.
И в естественном, и во вторичном ареале индивидуальная изменчивость и качественных, и количественных признаков семян E. lobata выражена слабо. Однако эти признаки сильно варьируют на внутри — и межпопуляционном уровнях, хотя клинальной изменчивости признаков в инвазионных популяциях не прослеживается. Выявлено 17 дискретных вариаций семян, встречающихся и в естественном, и во вторичном ареале [Виноградова, 2007]. Степень реализации фенофонда в 90% изученных популяций составляла 100%; лишь в двух популяциях на северном пределе вторичного ареала отсутствовало 2-3 фена. Таким образом, в инвазионных популяциях перекрёстноопыляющихся видов, расширяющих вторичный ареал (к которым, несомненно, относится E. lobata), сохраняется высокий уровень генетического разнообразия, и именно это обеспечивает успешную адаптацию видов к новым условиям произрастания.
Интродукционные популяции закладывали неоднократно из семян, собранных от Закарпатья до Владивостока и от Вологды до Таганрога; всего испытаны 43 локальные популяции.
а) Изменчивость периода покоя семян. При весенних сроках посева не отмечено заметных отличий в темпах прорастания как нестратифицированных, так и стратифицированных семян различного географического происхождения: семена начинали прорастать на 5-6-е сутки после посева и полностью прорастали на 10-й день. Однако при осеннем посеве проявляется тенденция к более быстрому прорастанию семян, собранных в северной части ареала. Например, в 1984 году к 18 апреля взошло 26% семян северного происхождения (из Ленинградской, Новгородской и Вологодской областей), 15% семян московского происхождения и только 8% семян закарпатского происхождения.
б) Изменчивость фенологического ритма развития и темпов роста. В 1978 году растения московского и закарпатского происхождения вступили в генеративный период развития на 1,5 мес. раньше, чем владивостокского, причём последние к концу вегетационного сезона не успели образовать полноценные всхожие семена. Интенсивность и продолжительность роста убывали в следующей последовательности: владивостокские образцы (длина главной оси в конце вегетации 2,5-3 м), московские и закарпатские (1-2 м). Разница в сроках вступления в одну и ту же фенофазу между московскими и закарпатскими экземплярами составляла всего 1 неделю, а по длине главной оси образцы достоверно не различались. Это дало нам основание утверждать [Виноградова, 1992], что внутри восточноевропейской части ареала адаптивная изменчивость биологических признаков эхиноцистиса чётко не выражена, а имеются лишь два биотипа, отличающихся по темпам роста и фенологическому ритму развития — восточноевропейский и дальневосточный.
Суровая зима 1978/79 наложила серьёзный отпечаток на распространение эхиноцистиса. Многие из наблюдавшихся нами популяций вымерзли совсем, а оставшиеся насчитывали всего по 3-5 особей. Именно после этой зимы благодаря естественному отбору наиболее морозостойких экземпляров началось стремительное продвижение E. lobata в северные регионы.
Бурное расширение ареала сопровождалось значительным темпом адаптивной эволюции: эхиноцистис выработал и генетически закрепил важнейшие адаптивные признаки: более короткий период роста и быстрое прохождение полного цикла развития (на 2-3 месяца скорее) по направлению с севера на юг. В интродукционной популяции у самых северных новгородских образцов семена прорастают на 2-3 недели раньше, период роста заканчивается в конце июля, главная ось практически не ветвится, общая длина всех побегов не превышает 1-2 м, цветение наступает в середине июня, плоды завязываются в пазухе 5-13-го листа. У закарпатских образцов, напротив, период роста продолжается до первых заморозков, растения сильно ветвятся, образуя побеги II, III и даже IV порядка, общая длина всех побегов достигает 9-13 м, цветение наступает в конце августа, плоды завязываются в пазухе 60-80-го листа, семена практически не вызревают. Внутрипопуляционная изменчивость изученных адаптивных признаков значительно ниже межпопуляционной.

Фенология образцов E. lobata различного географического происхождения
Сумма эффективных температур к моменту начала той или иной фенофазы во все годы наблюдений различалась столь сильно, что мы констатируем отсутствие корреляции между этими величинами.
в) Изменчивость фенологии и темпов роста в ряду последовательных поколений. Сравнение фенологического ритма родительских и дочерних особей проводили путём одновременного высева Р- и F1 — семян. При свободном опылении в интродукционной популяции уже в 1-м поколении происходит сближение феноритмов дочерних особей как южных, так и северных образцов с образцами местного, московского происхождения: дочерние особи вологодского происхождения зацветают на неделю позднее родительских, а дочерние особи закарпатских образцов — на 2 недели раньше родительских. Однако по количественным признакам, характеризующим нарастание биомассы, родительские и дочерние экземпляры не различаются. Следовательно, рост растений — показатель менее пластичный, чем феноритм. При изменении условий существования растения в течение по меньшей мере 1 года сохраняют темпы прироста вегетативной массы, тогда как феноритм их изменяется в сторону сближения с феноритмом образцов местного происхождения.
г) Влияние длины дня. Поскольку мы не выявили зависимости между феноритмом растений и суммами эффективных температур, был поставлен опыт по выращиванию образцов различного географического происхождения при разных световых режимах. В середине июня, когда на растениях полностью раскрылся 1-й настоящий лист, опытные экземпляры накрывали плотной чёрной бумагой, создавая короткий 8-часовой световой день. Контрольные экземпляры росли при полном световом дне, который в июне на широте Москвы составляет 17,3 часа.
Многолетние наблюдения показали, что растения одного и того же места сбора зацветали в одно и то же время, несмотря на различия суммы эффективных температур к дате начала цветения.
У растений северных репродукций длина главной оси и число её метамеров выше в условиях короткого дня, у растений же южных репродукций, наоборот, длина главной оси и число её метамеров выше в условиях длинного дня, у московского образца достоверных различий между опытными и контрольными экземплярами не зафиксировано. Ритм развития растений в условиях короткого дня с севера на юг убыстряется практически в 2 раза. У растений северных репродукций феноритм контрольных и опытных экземпляров не различается; растения московской репродукции при коротком дне вступают в генеративный период развития на 2 недели раньше, а карпатской репродукции — на 1-1,5 месяца раньше, что даёт последним возможность на широте Москвы завязать плоды, а иногда и дать зрелые семена.
Сравнение морфометрических параметров образцов E. lobata из южной и северной части вторичного ареала, выросших в условиях длинного и короткого дня.
1 — в условиях длинного дня; 2 — в условиях короткого дня; а — длина главной оси E. lobata из Новгородской области (Едрово); в — длина главной оси E. lobata из Ужгорода; г — общая длина всех побегов E. lobata из Ужгорода
Таким образом, сформированный во вторичном ареале и генетически закреплённый феноритм E. lobata в значительной степени обусловлен продолжительностью длины дня в местах его произрастания.
Результаты изучения самоопылителей, апомиктов и вегетативно размножающихся видов
Chamomilla suaveolens появилась в Европе в середине XIX века и начала внедряться в нарушенные ценозы как «беженец» из ботанических садов. В южной Финляндии и в Петербурге этот процесс занял 20 лет, в Москве — 10 лет, в Томске — 4 года. Бурная натурализация вида наблюдалась в начале XX века. Следовательно, между первым появлением Ch. suaveolens в Европе и натурализацией вида прошло не менее 20 смен поколений. В настоящее время в России произрастает по меньшей мере 100-ое поколение этого вида.
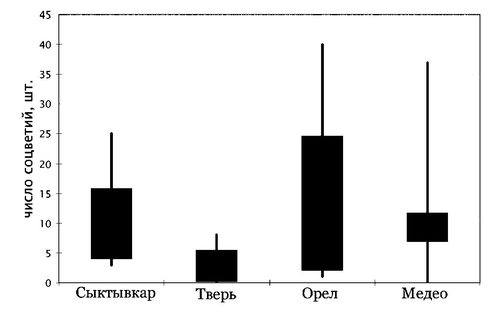
Изменчивость морфометрических признаков Ch. suaveolens изучали в интродукционной популяции, сформированной из семян, собранных в Сыктывкаре, Твери, Москве, Орле и в Казахстане в окрестностях Алма-Аты [Виноградова, 2001].
У образцов сыктывкарской и орловской репродукций цветоносные побеги начали появляться в середине июня, у растений алма-атинского происхождения — в конце июля, а у образца из Твери — только в середине августа. Экземпляры сыктывкарского происхождения зацвели в начале июля, орловского — на 2 недели позднее, алма-атинского — на месяц позднее и тверского на 2 месяца позднее. У образца из Твери 25% растений так и не образовали ни одного цветоносного побега и погибли от осенних заморозков на стадии розетки.
Период роста сыктывкарских растений закончился к началу августа, образцов из Орла и Медео — к началу сентября, а тверской образец продолжал медленно расти до начала октября. К концу вегетационного периода тверской образец имел самые короткие побеги, наименьшее число листьев и корзинок и достоверно отличался от всех остальных образцов. Отмечено и достоверное различие по длине побега между сыктывкарскими и орловскими образцами.
Изменчивость количественных признаков образцов Chamomilla suaveolens в интродукционной популяции
Таким образом, у ромашки душистой в её вторичном ареале выявлено наличие нескольких биотипов, различающихся по фенологическому ритму развития. При этом изменчивость феноритмов носит неупорядоченный, а не клинальный характер.
Bidens frondosa в естественном ареале является высокополиморфным видом. Различные разновидности В.frondosa заносились в Европу с 1762 года, однако натурализовались только две из них: В. frondosa var. frondosa с тройчатосложными листьями, короткими листочками обертки и коричневыми бородавчатыми семянками и B. frondosa var. anomala, которая отличается от типичной разновидности вверх направленными зубцами на ости семянки. Началом формирования вторичного ареала B. frondosa var. frondosa в Европе следует считать 1896 год, a B.frondosa var. anomala — 1954-й.
На территории бывшего СССР существует три изолированных очага распространения B. frondosa. Приморье, куда она была занесена из Америки не позднее 1950 года, запад европейской части, где этот вид стал расселяться с 1955 года, и Черноморское побережье Кавказа, куда череда занесена в конце 1970-х годов. Все растения представляют собой типичную разновидность. Единственная находка B. frondosa var. anomala сделана в 2003 году в Москве. За последние 15-20 лет B. frondosa начала активно расширять вторичный ареал и внедряться в естественные фитоценозы по берегам водоёмов, вытесняя аборигенную B. tripartita [Виноградова, 2009].
Интродукционная популяция заложена из семян приморского (окрестности озера Ханка), ростовского (Ростов-на-Дону) и московского происхождения [Виноградова, 2003]. Экземпляры, выросшие из дальневосточных семян, начали бутонизировать в середине сентября и погибли от осенних заморозков, так и не начав цвести. Экземпляры московского происхождения образовали бутоны в середине июля, зацвели в начале августа и дали плоды в конце сентября.
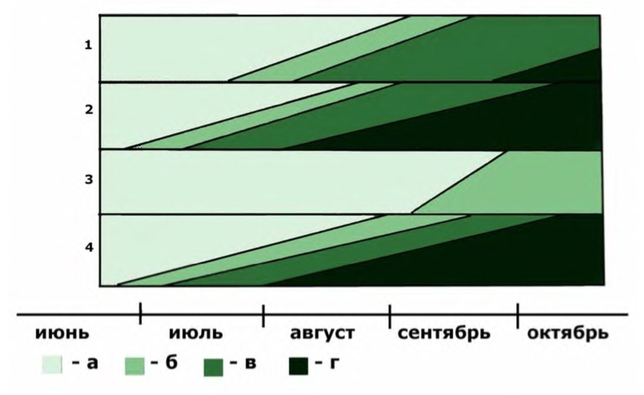
Феноритм Bidens frondosa в интродукционной популяции при различной длине дня
1 — образцы московского происхождения, естественная длина дня; 2 — короткий день; 3 — образцы из Приморья, естественная длина дня; 4 — короткий день; а — вегетация; б — бутонизация; в — цветение; г — плодоношение.
Отмечен более быстрый рост растений московского происхождения, максимальная интенсивность которого приходится на июль. Максимальная интенсивность роста дальневосточных образцов приходится на август. Поэтому в начале августа московские экземпляры были значительно выше дальневосточных и имели больше метамеров, а в начале октября оба образца по высоте уже не различались, а по числу метамеров дальневосточные растения даже достоверно перегнали московские.
Внутрипопуляционная изменчивость ритма роста и фенологии B. frondosa оказалась очень низкой. Коэффициент вариации даже самого изменчивого признака — высоты растений — составлял всего 8-12%.
Влияние длины дня на рост и фенологию череды в интродукционной популяции изучали путём сравнения темпов роста и феноритмов опытных растений, произраставших в стадии проростков в течение 3 недель при искусственном коротком дне (9 часов) с контрольными растениями, произраставшими при естественной длине дня [Майтулина, 1990]. Опытные растения и московского, и дальневосточного происхождения начали бутонизацию в конце июня и полностью прошли весь жизненный цикл до середины октября. Таким образом, при коротком дне московские образцы вступают в генеративный период развития на месяц раньше контрольных, а дальневосточные — на 2,5 месяца раньше контрольных, что даёт им возможность сформировать на широте Москвы зрелые семена. По темпу роста опытные растения различного географического происхождения друг от друга не отличались. Следовательно, в дальневосточной части ареала B. frondosa является короткодневным растением и на широте Москвы обнаруживает более медленный рост и неспособность вступить в стадию цветения. Наоборот, в европейской части ареала произрастает длиннодневная форма череды, отличающаяся более быстрым ростом и способностью полностью проходить жизненный цикл при естественной длине дня на широте Москвы.
Между образцами, выросшими из семян, собранных внутри изолированных очагов вторичного ареала, не отмечено различий ни по темпам роста, ни по фенологическому ритму развития.
Impatiens parviflora появилась в Европе в 1830-е годы, начала дичать через 10-15 лет, стремительное расширение её ареала и натурализация начались через 120 лет. В Европейской России и Балтии культивировалась с 1870-х годов, дичать стала через 50 лет, превратилась в массовый сорняк через 100 лет.
Интродукционная популяция закладывалась и при осеннем, и при весеннем посеве семян [Майтулина, 1988; Виноградова, 1992]. Семена жёлто-цветковой формы собраны в Санкт-Петербурге (4 образца), Вологде (1 образец), Москве (2 образца, один из них имел почти белые лепестки), Алма-Ате (1 образец), Таджикистане (3 образца с Памира) и Туркмении (1 образец с хребта Кугитанг); семена лилово-цветковой формы собраны в Туркмении (1 образец с Кугитанга) и Таджикистане (6 образцов).
При осеннем посеве единичные всходы появились во второй декаде, а массовые всходы — в третьей декаде апреля. Практически одновременно (в 20-х числах мая) растения вступили в фазу бутонизации, в первой декаде июня — в фазу цветения. В середине июня все растения образовали плоды, а в начале июля стали разбрасывать семена. Исключение составили жёлто-цветковые образцы из Рушана и Кугитанга, у которых долгое время часть особей не вступала в стадию цветения, а также лилово-цветковые образцы из Комсомолабада и Калаи-Хумба, плоды которых стали быстро растрескиваться и рассеивать семена. Однако эти различия не превышали 2-х недель. Следует отметить, что у особей, находящихся в стадии плодоношения и семеношения, новые бутоны и цветки продолжали образовываться вплоть до осенних заморозков.
Уже в середине июля после семеношения часть особей стала засыхать, но только лилово-цветковый образец из Кугитанга полностью выпал в конце сентября. У остальных образцов часть особей отмирали, а другие продолжали развиваться до середины октября.
Амплитуда изменчивости высоты растений была невысокой. Имели место достоверные межпопуляционные различия между образцами по этому параметру, но они не носили клинального характера.
Число метамеров главного побега, число боковых побегов, цветков и плодов оказались весьма изменчивыми как внутри образца, так и между образцами.
При весеннем посеве нестратифицированными семенами к середине мая проросли лишь единичные экземпляры из Кугитанга и Таджикистана, которые начали бутонизировать в середине июня. Следовательно, в естественном ареале имеются отдельные особи, не нуждающиеся в стратификации семян.
Таким образом, у недотроги мелкоцветковой наблюдается сильная внутри-популяционная изменчивость по числу метамеров побега, числу цветков и плодов и по степени ветвления, сильная межпопуляционная изменчивость (не носящая, однако, закономерного характера) по высоте, числу метамеров главного побега, числу цветков и плодов и по степени ветвления. Существенных различий по темпам роста и фенологии как внутри образцов, так и между образцами, не отмечено.
Amaranthus albus в 1860-х годах был занесён в порты Чёрного моря и начал распространяться по южным районам Российской империи. Однако в ботанических садах Москвы и Санкт-Петербурга его культивировали ещё в 1812 году [Виноградова, 2002].
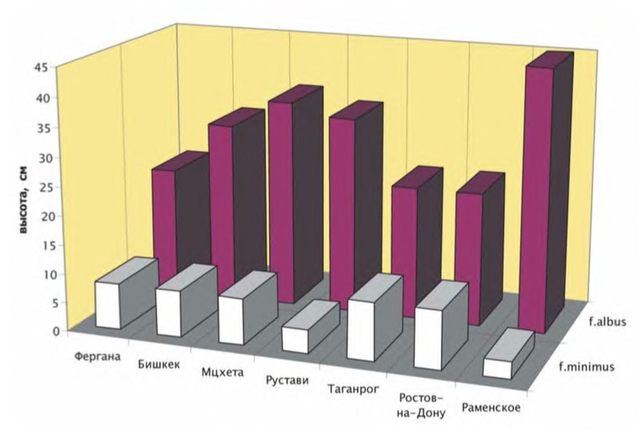
При осеннем посеве семян, собранных в Московской области, Узбекистане, Абхазии и Киргизии, всходы у всех образцов появились в последней декаде мая. Растения дружно прошли цикл развития и дали зрелые семена: образцы из Ферганы, Сухуми и Подмосковья — в конце июля, а образец из Бишкека — на неделю позднее. Никаких других различий между образцами не наблюдалось — все представляли собой типичную форму A. albus f. albus. Более того, количественные признаки (высота, число побегов II и III порядков, размеры листовой пластинки) также варьировали незначительно.
При весеннем посеве нестратифицированных семян, собранных в Московской и Ростовской областях, Узбекистане, Абхазии, Грузии и Киргизии, всходы появились через 12 дней после посева, цветение наступило через 40 дней после прорастания и продолжалось до осенних заморозков, семена образовались через месяц и стали осыпаться через 1,5 месяца после начала цветения. Все эти «весеннепроросшие» растения образовали побеги II, а некоторые — и III порядка; ни стелющихся, ни подушкообразных особей отмечено не было. Никаких различий ни в морфологии, ни в сроках прохождения фенофаз нами не выявлено — все особи относились к типичной A. albus f. albus. Наибольшей высотой отличался подмосковный образец, он же содержал наибольшее число особей (45%), имеющих побеги III порядка. Самыми низкими и слабоветвящимися оказались оба образца из Ростовской области (средняя высота 23 см).
Средняя высота двух форм Amaranthus albus в интродукционной популяции
В июне — начале июля начали прорастать оставшиеся в почве семена всех образцов. Выросшие из них «летнепроросшие» неотеничные особи скорее вступили в генеративный период развития, чем весенние, но росли очень медленно, в конце вегетации не превышали 10 см в высоту, совершенно не ветвились и представляли собой A. albus f. minimus. К осени образцы из Средней Азии на 22-26% состояли из A. albus f. minimus, образец подмосковного происхождения — на 33%, грузинского — на 44-62%, а в образцах ростовского происхождения эти карликовые экземпляры превалировали и составляли от 46 до 75%. Наименьшая высота экземпляров A. albus f. minimus отмечена в подмосковном образце (средняя высота 3 см) и образце из Рустави (4 см). У остальных образцов высота A. albus f. minimus составляла 8-10 см.
Таким образом, описываемые в литературе формы A. albus являются отражением модификационной изменчивости вида. Генетически закреплены лишь две формы: типичная A. albus f. albus, семена которой прорастают весной, и неотеничная A. albus f. minimus, семена которой прорастают в середине лета. Общая генетическая изменчивость A. albus во вторичном ареале очень низка, что объясняется обеднённым генофондом первоначальных малочисленных инвазионных популяций в сочетании с самоопыляемостью вида. При этом изменчивость феноритмов не выражена, а изменчивость ритма роста носит неупорядоченный характер.
Epilobium adenocaulon занесен в северо-западные районы СССР в конце XIX века, однако встречался крайне редко вплоть до 1960-х годов [Скворцов, 1976]. При осеннем посеве семян, собранных в Архангельске, Москве, Сыктывкаре и Вологде, не отмечено достоверных различий между образцами ни по темпам роста, ни по фенологическому ритму развития. Растения зацвели одновременно в конце июля, через 4 недели образовали семена и оставили зимующие розетки, которые на следующий год так же дружно вступили в генеративный период развития и погибли.
При осеннем посеве из большего числа пунктов сбора семян — от Ростова-на-Дону до Вологды — различий в феноритме также не наблюдалось. Однако замечены различия по высоте побегов: чем севернее находилось место сбора семян, тем выше были растения. Исключение составили экземпляры тульской репродукции, имеющие такую же среднюю высоту, что и растения из Ростова-на-Дону. Высота ростовских растений варьировала от 6 до 53 см (в среднем 30 см), липецких — от 7 до 59 см (в среднем 37 см), тульских — от 10 до 66 см (в среднем 30 см), московских — от 28 до 72 см (в среднем 46 см), вологодских — от 22 до 62 см (в среднем 49 см).
Таким образом, во вторичном ареале в Европе E. adenocaulon имеет очень низкую генетическую вариабельность. Различий по феноритму не наблюдается, а изменчивость ритма роста выражена недостаточно отчетливо.
Galinsoga parviflora интродуцирована в Англию в 1796 году, найдена там вне ботанического сада через 13 лет; массовое ее распространение началось только через 125 лет! В Дерптском и Санкт-Петербурском ботанических садах G.parviflora стала расширять свой ареал лишь через 135 лет! Поскольку этот вид является однолетником, можно считать, что период его натурализации на территории бывшего СССР насчитывает не менее 50 смен поколений.
Интродукционные популяции этого вида закладывали неоднократно путем как осеннего, так и весеннего посева из семян, собранных в 1981 году в Санкт-Петербурге и Москве; в 1983 году в Санкт-Петербурге и Таджикистане (Хорог, Рушан); в 1985 году в Москве и Узбекистане (Коканд, Фергана); в 1987 году на Украине (Львов, Славяногорск), в Армении (Эчмиадзин), Грузии (Тбилиси) и в Киргизии (Бишкек). При любых вариантах образцы практически не различались по темпу роста и фенологии. Выращивание образцов при различной длине дня также не оказало влияния на феноритмы: и опытные, и контрольные экземпляры изучаемых образцов начали цвести в конце июля, а плодоносить — в конце сентября [Виноградова, 2002].
Следовательно, популяции G.parviflora на всем протяжении вторичного ареала в нашей стране являются гомогенными по важнейшим адаптивным признакам: ритму роста и фенологическому ритму развития. Однако наблюдается значительная межпопуляционная изменчивость по степени опушенности верхней части стебля, цветоложа и обертки.
Galinsoga quadriradiata культивировалась в ботаническом саду Санкт-Петербурга с 1842 года. В одичавшем виде стала встречаться в Европейской России с 1920-х годов. Массовое распространение на Украине и в Белоруссии началось в послевоенные годы, а в центральных и северо-западных районах России и в Средней Азии — в 1970-е годы. Поэтому можно считать, что в настоящее время в Восточной Европе и Средней Азии произрастает по меньшей мере 40-е поколение вида.
Интродукционные популяции закладывали неоднократно путем как осеннего, так и весеннего посева из семян, собранных в 1982 году в Вологде, Москве и Таганроге; в 1983 году в Санкт-Петербурге и Душанбе; в 1985 году в Москве, Узбекистане и Киргизии; в 1987 году на Западной Украине и в Грузии [Виноградова, 2003].
При весеннем посеве стратифицированными семенами разрыв между началом цветения северных и южных образцов составляет всего 5 дней. При осеннем посеве наиболее заметные различия в феноритме отмечены в 1985 году: образец из Киргизии начал цвести на 2 недели позднее остальных образцов. Других вариаций по ритму роста и фенологическому ритму развития не отмечено.
Acorus calamus интродуцирован в Европу в XVI веке. Период произрастания инвазионных популяций аира в России насчитывает, по-видимому, более 300 лет.
Интродукционная популяция сформирована путем посадки фрагментов корневищ, собранных во Пскове, в Московской области, Армении, Грузии, Эстонии, Таганроге и Хабаровске [Виноградова, 2004].
В середине апреля начинают развертываться 3-4 весенних листа, которые постепенно отмирают к середине мая и окончательно засыхают в середине июня, достигнув средней длиы от 6-7 (Таганрог и Псков) до 11 см (закавказские и подмосковные образцы). Коэффициент вариации длины весенних листьев очень высок (не ниже 83%)!
Одновременно с отмиранием весенних листьев начинают отрастать летние листья, развертывание которых завершается в конце июля. У подмосковных и таганрогских образцов образование летних листьев начинается в первой декаде мая, у остальных на 1-1,5 недели позднее. За вегетационный период на одном растении образуется 6-7 летних листьев, длина которых составляет в среднем от 50 (Псков) до 114 см (Хабаровск). Различия по длине летних листьев практически между всеми образцами достоверны; коэффициент вариации этого признака — от 6% (Тарту) до 60% (Псков).
Летние листья у растений закавказского происхождения начали засыхать на 2 недели раньше, чем у остальных, более северных, образцов. Полное отмирание летних листьев отмечено в начале сентября. В середине августа начинают расти листья (по 4-5 на особь), которые уходят под снег и в следующем вегетационном сезоне развиваются как весенние листья.
У подмосковных и таганрогского образцов соцветия появились в первой декаде июня, остальные образцы зацвели на 1-1,5 недели позднее. На каждом растении образовалось только по 1 початку, средняя продолжительность цветения составляла 3 недели. Длина покрывала всегда превышает длину листьев и составляет в среднем от 73 (Грузия) до 133 см (Подмосковье). Длина соцветия у всех образцов 6-8 см, диаметр 1,1 см. Зрелых семян после отцветания не обнаружено.
Таким образом, во вторичном ареале аира на территории европейской части России и Закавказья выявлена генетически закрепленная межпопуляционная изменчивость (не носящая клинального характера) по темпам роста как весенних, так и летних листьев, однако изменчивость фенологического ритма развития практически не выражена.
Заключение
Изученные виды являются наиболее яркими представителями видов-ко-лонизаторов. Они имеют обширный вторичный ареал (в большинстве случаев значительно превышающий площадь естественного ареала), в котором наблюдается более широкая амплитуда изменчивости природно-климатических факторов. Все эти виды на родине произрастают вне зоны экологического оптимума, а вследствие этого обладают довольно широкой нормой реакции. Ни один из видов, доминирующих в естественных ценозах, не стал инвазионным!!!
Общей чертой натурализующихся видов является относительная генетическая бедность инвазионных популяций при широкой норме реакции. Относительное генетическое обеднение обусловлено происхождением инициальной популяции от одного или нескольких семян или кусочков побегов. Поэтому между появлением вида на новой территории и его внедрением в естественные ценозы проходит длительное время — период накопления микромутаций и рекомбинаций, продолжающийся у некоторых видов до 100 смен поколений.
Высокая фенотипическая пластичность позволяет генетически однородным популяциям приспосабливаться к различным местообитаниям. Вследствие формирующего эффекта, вызванного различием местообитаний, идёт генетическое дифференцирование популяций и наступает период бурного, иногда взрывного, расселения, поэтому к прогнозам относительно опасности или безопасности того или иного нового иммигранта относиться следует с превеликой осторожностью.
При расширении ареала период между первым появлением вида на новой территории и началом его натурализации стремительно сокращается и может продолжаться всего 3 года.
В популяциях натурализовавшегося вида формируются ряды изменчивости неадаптивных морфологических признаков. Амплитуда изменчивости этих признаков тем выше, чем дольше произрастает популяция в новых условиях. Так, в России полиморфизм Impatiens glandulifera ниже, чем в Западной Европе, a I. parviflora и Anisantha tectomm — выше, поскольку миграция первого вида шла в Евразии с запада на восток, а двух других — с юго-востока на запад. Амплитуда изменчивости зависит не столько от длительности периода натурализации, сколько от биологических свойств вида, из которых наиболее важными являются способ размножения и быстрота смен поколений.
Способ размножения не оказывает существенного влияния на натурализа-ционные возможности вида: к успешным колонизаторам относятся и размножающиеся вегетативно-подвижно Acorns и Elodea, и перекрёстноопылители Acer negundo и Impatiens glandidifera, и самоопыляющиеся виды рода Galinsoga. Однако способ размножения определяет механизм адаптаций. У перекрёстно-опылителей адаптация осуществляется путём перекомбинации генов и последующего естественного отбора, а у самоопылителей и апомиктов наиболее вероятны возникновение уже в инициальной популяции микромутаций физиологических признаков, связанных с расширением нормы реакции, и сохранение их в чистых линиях, способных к последующей натурализации. В первом случае имеет место движущий отбор, а во втором — стабилизирующий.
Особенности микроэволюции инвазионных видов
| Амфимикты | Вегетативно размножающиеся виды, самоопылители и апомикты | |
|---|---|---|
| Вид | Acer negundo, Echinocystis lobata, Impatiens glandulifera | Acorus calamus, Amaranthus albus, Ribes aureum, Aronia melanocarpa, Impatiens parviflora, Epilobium rubescens, Galinsoga parviflora, G. quadriradiata |
| Способ адаптации | Адаптация осуществляется путём перекомбинации генов и последующего естественного отбора | В инициальной популяции возникают микромутации физиологических признаков, связанные с расширением нормы реакции, которые сохраняются в чистых линиях, способных к последующей натурализации |
| Форма естественного отбора | Движущий отбор | Стабилизирующий отбор |
| Характер внутривидовой изменчивости | В ходе натурализации вырабатываются и генетически закрепляются важнейшие адаптивные признаки: менее продолжительный период роста и более быстрое прохождение полного цикла развития с юга на север. У Acer negundo с юга на север увеличивается также продолжительность периода покоя семян и морозостойкость. При этом наибольшее влияние на ритм роста и развития у растений длинного дня оказывают температурные условия и режим осадков, а у растений короткого дня — длина дня. | Во вторичном ареале наблюдается относительная константность биоморфологических признаков. Изменчивость ритма роста и развития либо практически не выражена, либо имеет беспорядочный межпопуляционный (не клинальный) характер |
У перекрестноопылителей во вторичном ареале формируется клинальная изменчивость фенологического ритма развития и ритма роста. В ходе натурализации вырабатываются и генетически закрепляются важнейшие адаптивные признаки: менее продолжительный период роста и более быстрое прохождение полного цикла развития с юга на север, что соответствует изменениям климатических условий вдоль широтного профиля. При этом в комплексе климатических факторов наибольшее влияние на ритм роста и развития у растений длинного дня оказывают температурные условия и режим осадков, а у растений короткого дня — длина дня.
У самоопыляющихся, апомиктных и вегетативно размножающихся видов в их вторичном ареале наблюдается относительная константность биоморфологических признаков. Изменчивость ритма роста и развития либо практически не выражена, либо имеет неупорядоченный межпопуляционный (не клинальный) характер, и это связано с тем, что отбор политолерантных линий или клонов происходит на самых ранних этапах натурализации, а далее они уже генетически почти не изменяются.
Во вторичном ареале могут иметь место и весьма значительные и константные, сопоставимые с уровнем видообразования морфофизиологические перестройки растений. Именно таков механизм возникновения Aronia mitschurinii.
← Предыдущая глава





