Золотая розга гигантская, или Золотарник гигантский
Solidago gigantea Ait.
Синонимы
Aster latissimifolius (Miller) Kuntze var. serotinus Kuntze; Doria dumetorum (Lunell) Lunell; D. pitcheri (Nuttall) Lunell; Solidago cleliae DC.; S. dumetorum Lunell; S. gigantea var. leiophylla Fernald; S. gigantea var. pitcheri (Nuttall) Shinners; S. gigantea subsp. serotina (Kuntze) McNeill; S. gigantea var. serotina (Kuntze) Cronq.; S. gigantea var. shinnersii Beaudry; S. pitcheri Nuttall; S. serotina Ait., 1789, not Retz. 1781; S. serotina var. gigantea (Ait.) A. Gray; S. serotina var. minor Hooker; S. serotinoides A.Love et D. Love; S. shinnersii (Beaudry) Beaudry; S. somesii Rydb.
Морфологические признаки
Solidago gigantea — высоковариабельный вид с неясным и трудным для оценки таксономическим статусом. Это прямостоячее многолетнее длинно-корневищное растение. Надземные побеги достигают от 30 до 280 см в высоту и от 5 до 11 мм в диаметре. Стебли ветвятся только в соцветиях, которые составляют в среднем 1/3 общей высоты побега, хотя такое соотношение довольно сильно варьирует (10-60%) и зависит от популяции, географического положения и времени цветения. Более высокие растения имеют тенденцию к позднему сроку цветения. Побеги до начала соцветия гладкие, часто пурпурные и покрыты беловатым восковым налётом.
Листья S. gigantea простые, очередные, продолговато-ланцетные, длиной 80-180 мм и шириной 10-30 мм. В течение вегетационного сезона образуется до 90 листьев. Более крупные листья образуются в середине стебля, листья мельче — в верхней и нижней частях стебля и в соцветиях. Листья с тремя более или менее заметными главными жилками; центральная жилка выделяется более явно, нежели латеральные жилки. Край листовой пластинки слабозубчатый вдоль всего листа, размер зубчиков варьирует в популяции весьма значительно. Листовые пластинки обычно гладкие сверху и снизу, но изредка могут быть опушёнными на нижней стороне по главной жилке.
В верхнем слое (10-20 см) почвы S. gigantea, в отличие от S. canadensis, образует длинные плагиотропные фиолетовые или красноватые корневища. Одно растение формирует за год от 3 до 50 корневищ. При неблагоприятном воздействии на растение число образующихся корневищ возрастает. Корневища достигают длины до 90 см и диаметра 1 см, часто ветвятся. На корневищах образуются почки (до 2,2 на 1 см), наибольшая их густота отмечается у основания стебля и на конце корневища.
Форма соцветий — пирамидальная метёлка. Обычно серповидно изогнутые веточки содержат многочисленные (1 200-190) корзинки [Schmid et al., 1988]. Корзинка диаметром 2-3 мм, обёртка соцветия имеет длину 3-4 мм. В корзинке имеется два типа цветков: краевые ярко-жёлтые язычковые женские и трубчатые обоеполые. Трубчатые цветки длиной 4-8 мм. Плоды длиной 1-1,8 мм с паппусом длиной 2,5-4 мм, опушённые длинными волосками, которые способствуют распространению семянок с помощью ветра [Weber, Jakobs, 2005].
От близкого вида, S. canadensis, хорошо отличается наличием корневищ, голым стеблем с беловатым налётом, неопушённой нижней стороной листа, более плотной архитектурой соцветия и коричневатым оттенком паппуса [Квитка, Виноградова, 2006].
Основное число хромосом Solidago gigantea х = 9, в естественном ареале у вида выделено три цитотипа: диплоид 2n = 18, тетраплоид 2n = 36, гексаплоид 2n = 54. J. Beaudry рассматривал цитотипы 2n = 18 и 2n = 36 как 2 отдельных вида: S. gigantea и S. serotina Ait. Позднее он выделил в пределах S. gigantea s. l. и третий цитотип — S. shinnersii, имеющий 2n = 54 [Melville, Morton,1982].
Ранее считалось [Weber, Schmid,1998], что в Европе натурализовалась только тетраплоидная форма S. gigantea. Однако в последнее время в Европе были обнаружены все три цитотипа [Weber, Jakobs, 2005], поэтому имеет смысл рассматривать S. gigantea как комплекс:
- S. gigantea 2n = 2х =18 (опушение по главной жилке с нижней стороны листа);
- S. serotina 2n = 4х =36 (гладкие узкие листья);
- S. shinnersii 2n = 6х = 54 (гладкие широкие листья).
Естественный ареал
S. gigantea естественно произрастает по всей Северной Америке южнее 55° северной широты, за исключением штата Аризона. Существует определённый характер в распределении трёх цитотипов: диплоидная форма тяготеет к востоку гор Аппалачи, тетраплоидная форма распространена по восточной лесной территории, гексаплоидная форма тяготеет к прериям.
Меры борьбы
S. gigantea, наряду с S. canadensis, внесена в «Список активно распространяющихся чужеродных видов Европейской и Средиземноморской организации по защите и карантину растений» [EPPO List...], в котором перечислены виды растений, которые представляют угрозу для благополучия растений, окружающей среды и биологического разнообразия на территории Европы.
В естественном ареале S. gigantea сильно повреждают тля, а также другие насекомые-вредители: Eurosta solidaginis, Rhopalomyia ssp., Trirhabda ssp. [Abrahamson et al., 1991, 2001], которые контролируют распространение данного вида. Напротив, во вторичном ареале у S. gigantea вредителей практически не наблюдается.
Имеется несколько механических способов удаления зарослей S. gigantea. Один из эффективных способов — кошение дважды в год (в мае и августе) в течение нескольких лет, или перекапывание почвы в течение лета при сухих погодных условиях. После сенокошения для контролирования роста зарослей S. gigantea следует высевать травосмесь, в результате чего значительно снизится густота побегов растений. Эффективные мероприятия по удалению плотных зарослей и подавлению жизнеспособности включают удаление раз в год с мульчированием, или удаление два раза в год без мульчирования в течение 3 летнего периода. Однако подобные механические воздействия могут нанести также вред естественной растительности, таким образом, способ этот рекомендовать нельзя. S. gigantea реагирует на удаление уменьшением диаметра стеблей и стимуляцией сильного повторного отрастания из корневищ. Кошение раз в год ненамного снижает густоту побегов и даже провоцирует противоположный эффект — увеличение плотности популяции на следующий год. Мульчирование подавляет рост сильнее, чем просто удаление, возможно потому, что срезанная трава содержит ингибиторы роста [Weber, Jakobs, 2005].
В Европе выясняют возможность препятствовать внедрению S. gigantea в леса посредством формирования вокруг них густых одновидовых зарослей аборигенных видов [Weber, Jakobs, 2005]. Контролировать расселение S. gigantea сложно и дорого. Искоренение вида нереально, и управление должно быть ограничено особыми площадями специальных экологических предприятий.
Обзор научных исследований и публикаций
Обнаружено [Schmid et al., 1988], что фотосинтетическая активность S. gigantea в начале вегетационного сезона высокая (19 ± 2 мmol CO2/m2/s), а позднее снижается (16 ± 2 мmol CO2/m2/s).
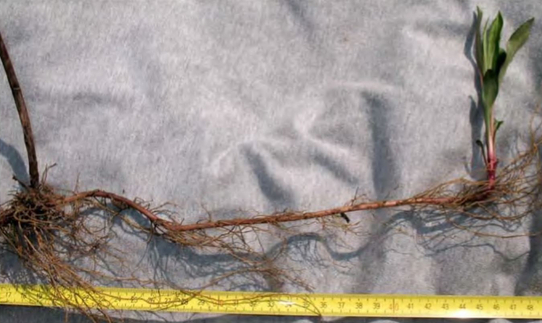
Корневища Solidago сanadensis (вверху) и Solidago gigantea (внизу)

Соцветия Solidago gigantea (слева) и Solidago canadensis (справа)
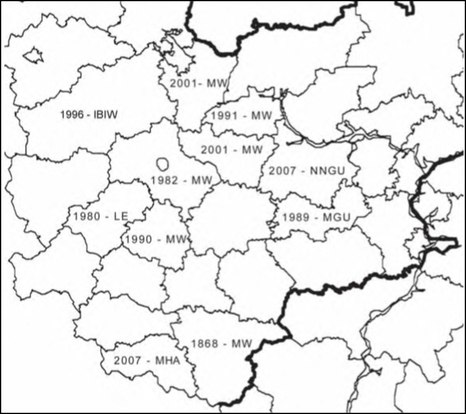
Расселение Solidago gigantea в Средней России

Внедрение Solidago gigantea в прибрежные сообщества

Золотая розга канадская, или Золотарник канадский
Solidago сanadensis L.

Дурнишник эльбский
Xanthium albinum (Widder) H.Scholz




