Костёр кровельный
Anisantha tectorum (L.) Nevski
Синонимы
Bromus tectorum L., Schedonorus tectorum (L.) Fries, Bromus setaceus Buckl., Zerna tectorum (L.) Lindm., Anisantha pontica C. Koch
Морфологические признаки
Озимый монокарпик или яровой однолетник. У яровых особей насчитывается от 1 до 5 побегов, у озимых — до 24 побегов. Длина побегов 20-50 см, число листьев на одном побеге варьирует от 3 до 7. Соцветие — односторонняя метёлка длиной 5-10 см с 14-25 тонкими, поникающими, дуговидно изогнутыми веточками.
В естественном ареале листья могут быть опушены смесью длинных и коротких волосков, только короткими волосками или, реже, быть голыми; в некоторых популяциях встречаются особи со всеми тремя вариациями опушения листьев. Во вторичном ареале отмечены только особи с опушёнными листьями.
Основание метёлки голое или опушённое; в долинных популяциях выше частота встречаемости неопушённого основания оси метёлки, а в высокогорных — опушённого основания [Виноградова, 2000а].
Нижняя цветковая чешуя длиной 20-30 мм, ланцетная или широко-ланцетная, с широким плёнчатым краем, наверху слегка притуплённая и с длинной остью. В естественном ареале преобладают растения, у которых верхняя цветковая чешуя полностью или хотя бы в нижней части голая. Ниже частота встречаемости растений с цветковой чешуёй, слабоопушённой короткими волосками; и очень редко встречаются единичные особи с цветковой чешуёй, сильно опушённой смесью коротких и длинных волосков. Во вторичном ареале преобладают особи с сильноопушенными цветковыми чешуями, а неопушённые цветковые чешуи вовсе не отмечены. Такая же закономерность прослеживается по степени опушения колосковых чешуй: в естественном ареале значительно преобладают особи с голыми колосковыми чешуями, а во вторичном ареале — с опушённой колосковой чешуей [Виноградова, 2000а]. В Северной Америке растения с колосками «скорее голыми, чем опушёнными» рассматриваются как A. tectorum var. glabratns Spenner. Встречается эта разновидность очень редко: в Канаде, например, найдена лишь в 4 пунктах в провинции Британская Колумбия, в 1 пункте в провинции Альберта и 1 экземпляр — в провинции Онтарио [Upadhyaya, 1986].
Наименьшее число цветков в колоске отмечено в естественном ареале у растений из равнинных популяций. У растений горных популяций среднее число цветков в колоске увеличивается до 4,1-4,4. Во вторичном ареале цветков в колоске ещё больше — до 5-8. Следовательно, на северном и высотном пределах ареала колосок состоит из большего числа цветков, что позволяет несколько увеличить число образовавшихся семян.
Во вторичном ареале в Волгоградской и Московской области встречаются целые популяции костра с побегами пурпурного цвета. В естественном ареале подобную окраску можно наблюдать только у единичных особей.
Естественный ареал
Родина A. tectorum — южные районы Восточной Европы и Западной Азии, Центральная Азия и Средиземноморье.
Меры борьбы
В посевах озимой пшеницы в Америке с костром кровельным борются путём весенней (март) и осенней (ноябрь) обработок посевов метрибузином в дозе 0,42 кг/га и метрибузином + тербутрином (0,28 + 0,67 кг/га). Лучшие результаты получены при осенней обработке метрибузином + тербутрином: сорняк уничтожается на 93%, а урожайность пшеницы повышается на 36% [Swan, Whitesides, 1988].
В качестве агента биологической борьбы с костром кровельным используется штамм D7 ризобактерии Psendomonas flnorescens [Kennedy et al., 2001].
Обзор научных исследований и публикаций
Отмечено, что при формировании вторичного ареала костра кровельного происходит снижение полиморфизма инвазионных популяций по признакам опушения. Некоторые аллели исчезают вовсе, а оставшиеся демонстрируют изменение частоты встречаемости (в естественном ареале преобладают особи с неопушёнными органами, а во вторичном с опушёнными). Проведено сравнение характера опушения верхней цветковой чешуи с данными L. Hulbert [1955], полученными на обширном материале, включающем 119 образцов из 21 пункта сбора в четырёх штатах США [Виноградова, 2006].
Отмечено значительное варьирование количественные признаки костра кровельного в естественных и инвазионных популяциях.
Сравнительная характеристика некоторых количественных признаков костра кровельного в естественном и вторичном ареалах
| Изученные признаки | Средние значения по всем популяциям | ||
|---|---|---|---|
| естественный ареал | вторичный ареал | ||
| Таджикистан | Россия | Северная Америка | |
| Число цветков на 1 растении | 292,5 (135-941) | 365,0 (238-531) | 327,5 [Mc Kone, 1985] |
| Число колосков на 1 растении | 81,3 (31-227) | 70,2 (52-91) | 108,5 [Mc Kone, 1985] |
| Число цветков в колоске | 3,6 (2-7) | 5,2 (4-8) | 3,0 [Mc Kone, 1985] |
| Длина верхней цветковой чешуи | 11,53 (10,7-14,40 | 12,52 (9,7-13,8) | 11,58 [Mc Kone, 1989] |
| Средняя высота, см | 26,3 (21,4-41,0) | 32,8 (23,5-52,0) | 70,4 [Mc Kone, 1985] 49,3 (35-65) [Hullbert, 1955] |
Имеются и другие данные, касающихся гетерогенности костра кровельного. Так, при исследовании изменчивости 19 польских популяций этого вида и 6 популяций из Португалии, Италии, Чехии, Словакии, Венгрии и Швеции [Krzakowa, Kraupe, 1981] в отношении систем ферментов установлено, что изученные популяции мономорфны по двум системам ферментов и полиморфны по двум другим системам ферментов эстеразе и глютамат-оксалат-трансаминазе.
Сходная работа [Novak, Mack, 1989] проводилась с использованием электрофореза в крахмальном геле для определения генетической вариабельности 24 популяций костра кровельного из его естественного ареала. Все популяции проанализированы на изоферментную вариабельность по 15 ферментам, кодируемым 25 генетическими локусами. Полученные результаты сравнили с данными, приводимыми ранее для североамериканских популяций. Обнаружена незначительная дифференциация по уровню полиморфизма внутри популяций костра кровельного и зафиксировано исчезновение некоторых редких аллелей при внедрении вида в Северную Америку. В евразийских популяциях 52% локусов были полиморфными, а в североамериканских только 28%.
Впоследствии по той же методике электрофореза в крахмальном геле был проведён [Novak, Mack, Soltis, 1991] анализ генетической вариабельности по 25 локусам 2 141 экземпляра костра кровельного, выращенных из семян, собранных в 60 популяциях в 4 регионах вторичного североамериканского ареала:
- восточная часть Скалистых гор (14 популяций);
- штаты Невада и Калифорния (14 популяций);
- Великая равнина (22 популяции);
- канадская провинция Британская Колумбия (10 популяций).
Общая генетическая изменчивость A. tectorum по сравнению с другими цветковыми растениями оказалась низкой. Полиморфность в пределах одной популяции составила 4,6%, охватывая 1,05 аллели на локус при средней ожидаемой гетерозиготности 0,012. Фактически гетерозиготность не выявлена ни у одной особи и реально составила 0,000. Генетическое сходство между популяциями равно 0,980. Авторы сделали вывод, что для A. tectorum, так же как для других самоопыляющихся видов, характерны низкая генетическая изменчивость и существенная генетическая дифференциация между популяциями.
В дальнейший анализ включено [Bartlett et al., 2002] 38 популяций A. tectorum из восточных штатов США для оценки их генетического родства с популяциями западных штатов. Генетическая изменчивость восточных популяций низкая: среднее число аллелей на локус — 1,01% полиморфных локусов в популяции — 1,05%, гетерозигот не обнаружено. Тринадцать популяций в восточной и западной части Северной Америки отличаются по локусу Pgm-1a и Pgm-2a, в то время как 8 популяций отличаются по локусу Mdh-2b и Mdh-3b. Однако несколько аллелей, обнаруженные в западной части Северной Америки (Got-4c, Got-4d и Pgi-2b), не были найдены в восточных популяциях. Следовательно, инвазия A. tectorum в Северную Америку произошло на обоих побережьях путём многократной интродукции. Исторические и генетические данные позволяют предположить, что восточные популяции возникли в результате как минимум двух независимых заносов. Распространение A. tectorum с востока на запад, вероятно, произошло в XIX столетии.

Метёлка Anisantha tectorum
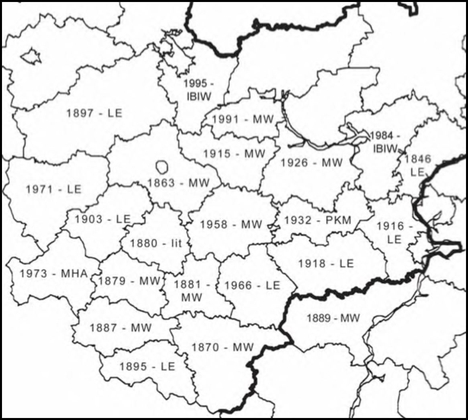
Расселение Anisantha tectorum в Средней России

Плодоношение Anisantha tectorum

Anisantha tectorum на ж/д насыпи, станции «Тарасовка» Московской области

Кислица прямостоячая
Oxalis stricta L.

Овсяница шершаволистная
Festuca trachyphylla (Hack.) Krajina




